|
In collaboration with the lab of Lars Behrendt, we are hiring a 2-year postdoc to work on developing a method to targeted cell-sorting and genomics of microbes. More information and how to apply here.
 Our lab is currently recruiting for an ERC Consolidator project on the origin of plastids in eukaryotes:
We are very excited for the publication of our habitat transition analysis to now be published in its final format in Nature Ecology and Evolution. The paper, which represent a major chunk of Mahwash's PhD, is available in full OA here. We used our approach of long-read metabarcoding (nearly 17K eukaryotic OTUs spanning the 18S-28S are presented) to reconstruct a phylogeny of eukaryotic diversity, and map onto it habitat transitions from marine to non-marine environments and vice versa (crossing the salt barrier). We also did ancestral state reconstruction to infer the habitat origin of the major eukaryotic groups. We show that species have successfully crossed the salt barrier more often that classically assumed, which likely was an important contributor to the diversification of life into what we can observe today.
We recently welcomed two new members in the lab:
But stay tuned for new openings: we will be recruiting two PhD students and two postdocs after the summer. We are incredibly honoured, humbled, and excited to receive an ERC Consolidator grant to pursue research on the origin of plastids. We are grateful to all the people who made this possible. Thanks to all, #ERC for the thorough review process, the 7 reviewers for their valuable feedback and the panel for their time and believing in this project. Stay tune for hiring announcement, we will hire PhD students and postdocs A fully funded PhD position is open in the lab, to work on figuring out what are the photosynthetic "organelles" in the protist Meringosphaera. The overarching goals of this four-year PhD project are to identify the closest free-living relatives to the endosymbionts, and to characterize the levels of cellular, genetic, and metabolic integration of the endosymbionts to pinpoint the nature of this endosymbiosis. A suite of single cell methods will be employed, including single-cell genomics and metagenomics, and we have also planned for confocal scanning microscopy, transmission electron microscopy, and focused ion beam-secondary electron microscopy (FIB-SEM). This is a collaboration with the lab of Rachel Foster at Stockholm University. More info and how to apply here: https://uu.varbi.com/en/what:job/jobID:464881/type:job/where:4/apply:1 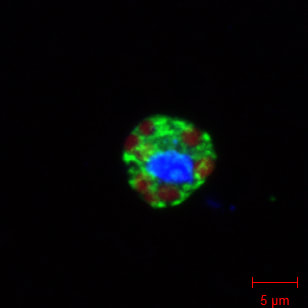 The editors at Nature Communications have chosen to feature our article “Single cell genomics reveals plastid-lacking Picozoa are close relatives of red algae” in the Editors’ Highlights webpage of recent research in “Ecology and evolution”. The Editors’ Highlights pages aim to showcase the 50 best papers recently published in an area. The highlights can be accessed at www.nature.com/collections/jadchbbhdi.  We are delighted that the Swedish Research Council has granted us funding for four years to continue our work on characterising the endosymbiotic relationship in the marine protist Meringosphaera. This work started off with a scholarship from Carl Tryggers that supported Dr. Vasily Zlatogursky (see here for the paper), and is now being carried out by Megan Sorensen in collaboration with the lab of Dr. Rachel Foster at Stockholm University with a postdoc grant from SciLifeLab. With this new grant, we will be seeking soon a PhD student; stay tuned.  |

